|
Quantitative morphogenesis
The overall behaviour of cells and their differentiation is clearly determined in a
predictable manner by
transcription factors and signalling pathways controlled by the genetic programme.
The behaviour of individual cells at a small time scale of seconds to minutes
is not genetically determined and predictable
but is controlled by self-organisation via interactions with their neighbours. These
interactions are assumed to involve mechanical processes. Biomechanics is a rather
new but fruitful topic in developmental and cell biology. Analysis is strongly based
on interdisciplinary approaches as quantification and modelling plays an important part.
Given the topicality
and interdisciplinarity
we are part in two major research initiatives, the DFG research unit
FOR1756
(Functional
dynamics of cell contacts in cellular assemblies and migratory cells) and the
SFB937
(Collective behaviour of soft and biological matter).
We established the following three projects:
-
Self-organisation of the nuclear array
Dynamics of cytoskeletal networks in syncytial embryos Junction dynamcis during cell intercalation
Self-organisation of the nuclear array
The early embryo represents a syncytium with many nuclei and
their associated cytoskeleton sharing a single big cell. As no separating
cell membranes are present, the cytoskeleton, such as microtubule asters
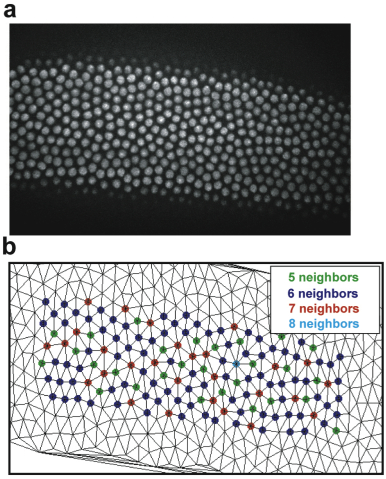
can directly interact. Tracking of nuclei showed that they are dynamic especially
after mitosis, when a doubled number of nuclei has to rearrange and find
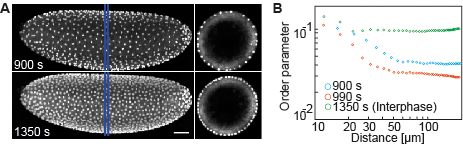
their new positions within a hexagonal array. We established a measure for nuclear
arrangement (order parameter) and
found that both, F-actin and microtubules, determine dynamics of the nuclear array.
We are now analysing the mechanism how forces are generated and transmitted between the
nuclei. Our hypothesis is that the four-headed motor and crosslinker Kinesin-5
is responsible for repulsion of adjacent microtubule asters and their associated nuclei.
A different role might be served by F-actin. We hypothesize that cortical actin dampens
movement of nuclei and microtubules by linking astral microtubules to the cortex.
(Kanesaki 2011, Koke 2014, funding: SFB937,
C Schmidt, Biophysics, T Aspelmeier, Math Stochastics,
U Schwarz, Theoret physics, Heidelberg))
Fluctuations of centrosomes
To comprehensively describe the dynamics of the cytoskeleton and nuclei
recording of their movement within the full frequency space from 1000 Hz to 0.01 Hz is required.
In the low frequency range with frame rates of few images per minute,
directional movements such as drift is uncovered. This has already been performed by conventional
live imaging. High frequency recording in the milli and micro second range would uncover
and passive (Brownian) movement. Most interesting is the range in between (sub second), as this may
reveal active non-equilibrium movements driven by the cytoskeleton.
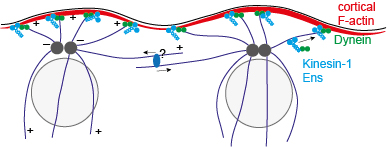
We established controsomal tracking at 1 Hz and analysed their fluctuations. We find that these
fluctuations are driven by microtubules and suppressed by cortical F-actin. Surprisingly, we found
that myosin-II does not contribute to active movement and that the microtubule motor Kinesin-1 dampens
centrosomal movement instead of promoting it as expected. As Kinesin-1 localises at the cortex and is required
for cortical localisation of Dynein, Kinesin-1 and Dynein may link the microtubule asters and the centrosomes
in their centre to the cortex.
We are now searching for the force generator and extend recording to the high frequency range with upto 1000 Hz.
(manuscr. in preparation, funding: SFB937,
Collaborator: C Schmidt, Biophysics, A Zippelius,
Theoret. Physics, T Aspelmeier, Math Stochastics)
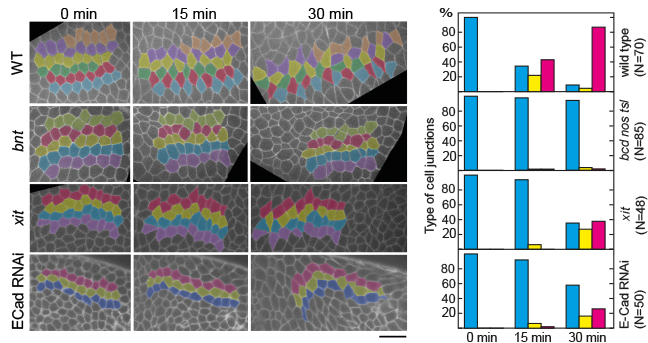
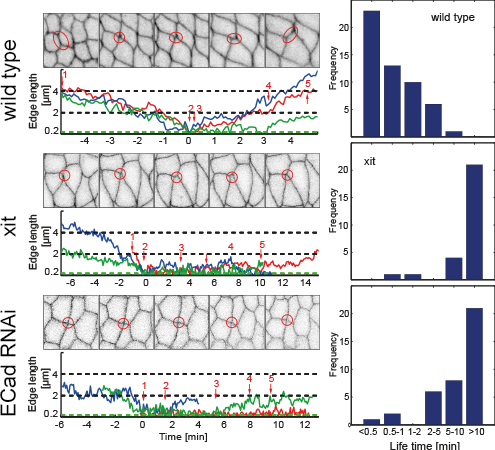
Dynamics of junctions in cell intercalation
Cell intercalation leads to tissue elongation that is central to many morphogenetic processes
during development. In epithelia, intercalation is driven by a concerted shrinking and
extension of cell junctions which can be described as a topological T1 process. It has
been previously proposed that junction dynamics is autonomously controlled by actomyosin
generated forces. We found that the exten- sion of new junctions is non-autonomous and
depends on the coordination of cell area changes be- tween the cells participating in the
T1 process. This coordination requires tissue tension and E-Cadherin. Large scale cuts in
the tissue lead to impaired resolution of 4x vertices. We propose to investigate the mechanism
of E-Cadherin dependent coordination in area changes. The coordinated cell dynamics will be
characterized using a high yield quantitative imaging approach developed in the initial
funding period. We will investigate candidates that may signal between adherens junctions
and actomyosin, such as Rho signalling and junction associated regulators of F-actin.
Furthermore, we will investigate the mechanism how tissue cuts affect the loss of coordination
in cell area changes. We will
test the hypothesis that E-Cadherin is the mediator of a mechanotransduction pathway that senses
the applied stress in one cell and elicits a signal in the other cell. We will establish methods
for the identification of stochastic dynamical systems models of intercellular coordination. These
models will enable us to accurately characterize the contribution of distinct junction-types to
intercellular coordination.

Exchange of neighbouring cells by junction dynamics (T1 process). A vertical junction constricts
so that two 3x vertices fuse into a single 4x vertex. The 4x vertex is resolved by
splitting into two 3x vertices. The new junction is in perpendicular orientation to the
old junction. The previous neighbours (blue) are not in contact with eachother anymore, whereas the green
cells are touching now.
(Zhang 2014, funding: DFG FOR1756,
Collaborator: F Wolf, MPI)
|