|
Epithelial morphogenesis and
Cell cycle switch
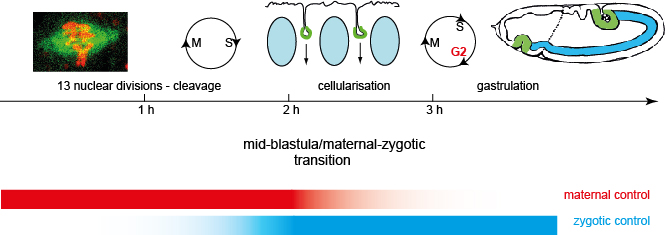
Drosophila embryonic development starts with a syncytial stage, in which the cell
cycle proceeds without cytokinesis. After about two hours, the cell cycle pauses in a G2 phase, allowing
the nuclei to be incorporated into cells. The process is called cellularisation and is unique
in the sense that an epithelium is generated de novo.At the end of cellularisation
the embryo consist of
a single layered uniform epithelium enclosing the interior yolk.
The transition from syncytial to cellular development marks the "mid-blastula
transition (MBT)". Concommitantly,
gene expression dramatically changes. Maternal RNAs are degraded. Prior to MBT only
maternally provided transcripts and protein are
present, as the zygotic genome has not been transcripted, yet.At and shortly before
MBT, RNA polymerase
is activated and zygotic genome is transcribed and consequently takes over control of
development and cell behaviour.
Due to the coupling of gene expression, cell cycle behaviour (pause in G2 phase), and
morphology change (cellularisation), MBT is regared as a paradigm for a developmental transition.
We address the following questions:
How are epithelial compartments established and maintained? How is actin dynamics controlled in the early embryo? What is the role of the slam RNA-protein complex in cellularisation? How is the cell cycle pause coordinated with cellularisation?
Formation of epithelial compartments and Actin dynamics
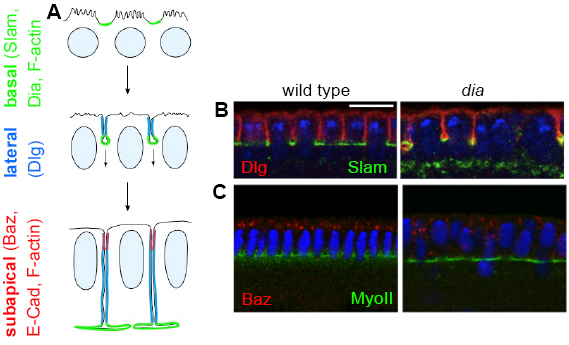
During cellularisation basal, lateral and sub-apical compartments are
established and in the following maintained.
We have previously identified a Slam-RhoGEF2-Rho1-Dia signalling cascade that restricts
Rho signalling and F-actin formation to the basal domain (furrow canal). We and others found that
F-actin is required for separation of lateral and basal domains but not for specification of the
basal domain. For example, the lateral marker Dlg is separated from basal markers (Slam, Patj, F-actin)
but spreads into the furrow canal in dia mutants, or embryos treated with Latrunculin. Separation
of epithelial domains may be achieved by (1) a physical barrier (e. g. a junction) at the interface,
(2) targeted vesicle transport or recycling, (3) differential stability, (4) differential diffusability.
As we observed that domains are separated in shi (dynamin) mutants and embryos without basal junctions (Sokacs 2008),
we currently favour the model that cortical F-actin restricts diffusion of lateral markers into the basal domain.
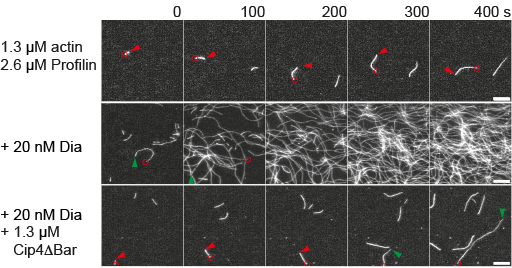
We have found genetic, biochemical and physical interactions of Dia and the F-Bar protein Cip4. Cip4 antagonises
Dia in vivo as well as in actin polymerisation assays with purified recombinant proteins. This was surprising,
as Cip4 is an activator for Arp2/3 dependent branched actin polymerisation.
We are now analysing the biochemical mechanism of Dia-dependent actin nucleation and elongation by Cip4.
(Grosshans 2005, Wenzl 2010, Kanesaki 2013, Yan 2013, funding: DFG Priority programme
Actin nucleators,
Collaborators: J Faix, Hannover, S Bogdan Münster)
Slam RNA-protein particles
The gene slam is required for cellularisation. In slam mutants the furrow is
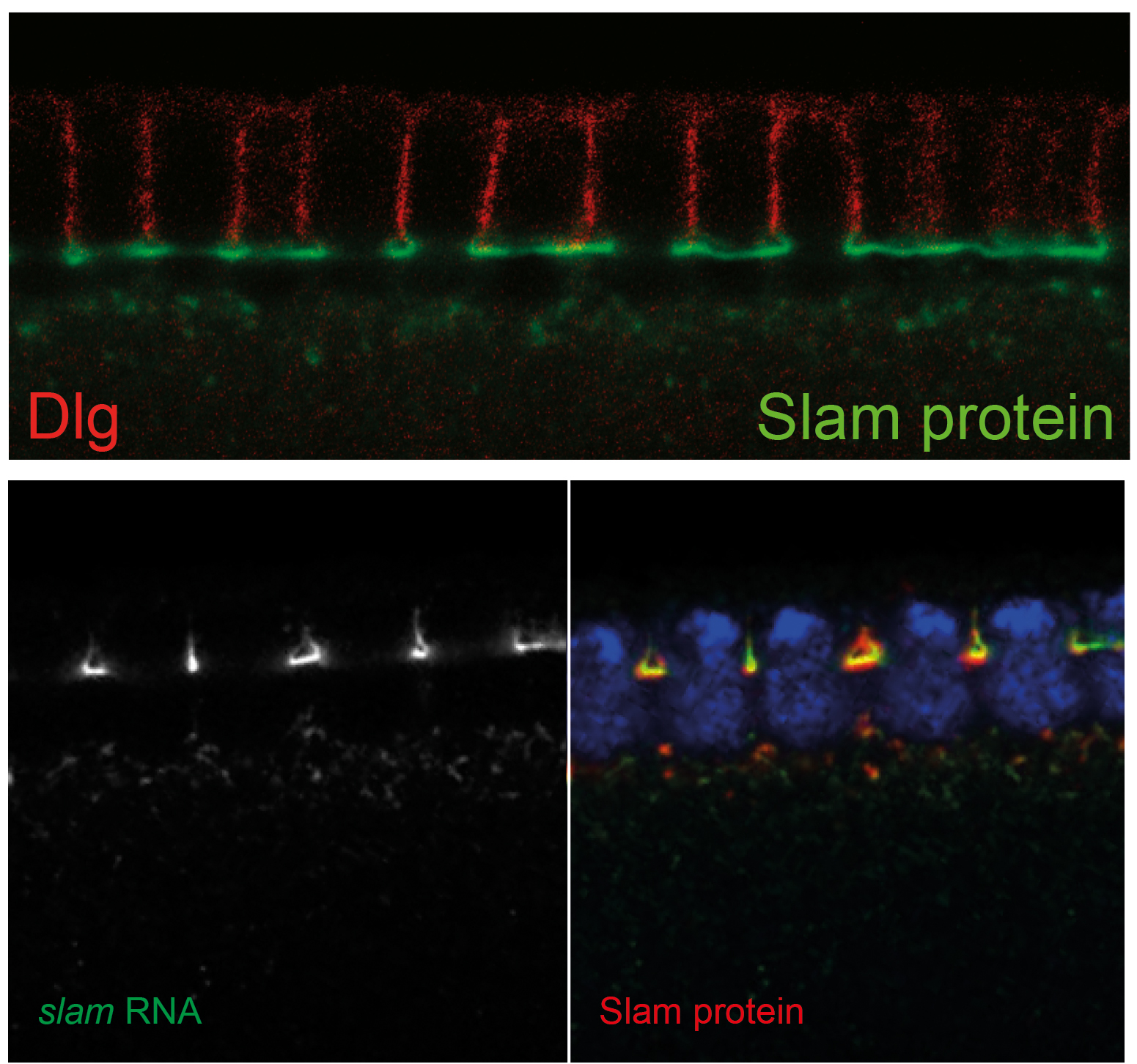
specified but does not invaginate. Slam protein strongly accumulates at the furrow canal
and thus correlates with its function. Surprisingly we found that slam mRNA also accumulates
at the furrow canal. Slam RNA and protein staining patterns almost match. Consitent with colocalisation
slam RNA and protein co-immunopreciptiate and bind in vitro.
We disected the mutual interaction of this unusual of mRNA and its encoded protein. We found that
Slam protein is required for mRNA localisation, thus representing a feed-back loop in the flow of
genetic information. Visa-versa mRNA localisation is required for slam protein expression, which represents
a non-coding function of the mRNA.
We are now studying the following two aspects:
1. What is the physiological function of this unusual mRNA-protein interaction?
2. What are the molecular determinants of this mRNA-protein interaction?
We have partially mapped the domains within the RNA and protein that are sufficient for interaction
and started to reconstitute the complex with the final aim of determining the structure of the RNA-protein complex.
The project is performed within the SFB860
(Integrative structural biology) , that provides the collaborations needed for structural analysis of
this novel RNA-protein complex.
(Wenzl 2010, Acharya 2014, funding: SFB860)
Cell cycle control
After fertilization, pronuclear fusion and completion of meiosis-II,
the nuclei proliferate in 13 rounds of synchronous cell cycles. In the following
interphase 14, the cell cycle mode switches. S phase is extended and the DNA is
replicated in an early and late replication. A long G2 phase is introduced for the first time.
We study the following two aspects:
1. What is the initial trigger for the switch of the cell cycle mode?
2. Which molecular mechanisms cause the introduction of the G2 phase?
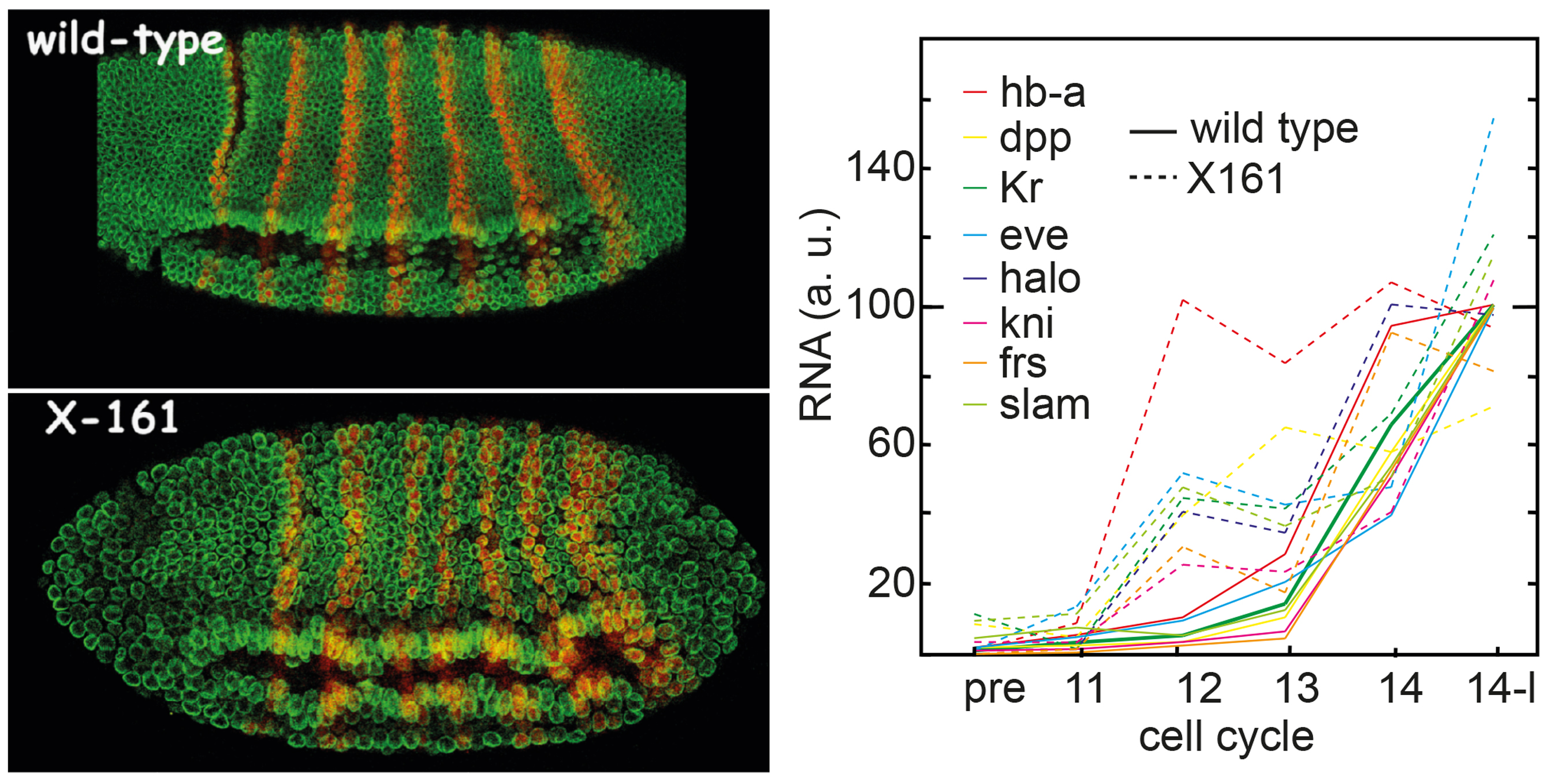
By analysis of the X161 mutation that undergoes a precocious switch including precocious onset of zygotic
gene expression,
we found that the timing of zygotic transcription
is the initial trigger for the cell cycle switch. We could reject other models that put
other processes also required for the switch, such as activation of the DNA replication/repair
checkpoint, maternal RNA degradation and reading of the nuclear-cytoplasmic ratio.
We now investigate the premature onset of gene expression comes about.
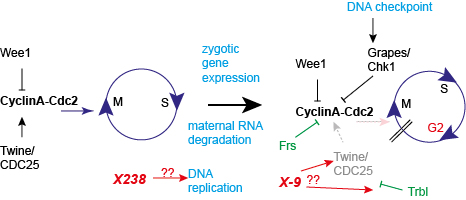
Secondly, we investigate how the G2 phase is introduced in interphase 14. The final readout
is inhibition of the Cdk1-CyclinA/B complex. We have previously shown that the zygotic gene frühstart
(frs)
is sufficient and partially necessary for the timely pause. We found that Frs binds to the hydrophobic
patch of Cyclin A and in this way inhibts Cdk1 kinase activity although the active center of the kinase
is not blocked by Frs binding. We hypothesize that only the phosphorylation of
a subset of Cdk1 substrates, i. e. the ones that
rely on binding to the hydrophobic patch, are inhibited.
A second mechanism is the induced degradation of the Cdc25 Twine, whose half-life time is reduced from about
20 min in previous cycles to only 2 min in interphase 14. Trbl is involved in destabilisation of Twine.
We are analysing mutations X238 and X9 that undergo a premature and delayed switch, respectively. Both genes
have been cloned by us. We are currently analysing how their proteins are involved in control of DNA replication
and degradation of Twine, respectively.
(Grosshans 2000, Grosshans 2003, Gawlinski 2007, Sung 2013)
|