Francisco Welter-Schultes
II. Zoologisches Institut der Universität
Berliner Str. 28
D-37073
Göttingen, Germany
E-mail:
Flying snails.
In this study, probable maximum flight distances for airborne transport of
loose specimens of Truncatellina land snails from Greece were calculated.
The results suggest that minute snails could be blown over the sea for several
kilometers in heavy storms.
Truncatellina is a minute land snail of the family Vertiginidae, not
exceeding 2 mm in size. In central Europe, T. cylindrica (same size)
is known quite well (click here for its range in Slovenia).
In 1995 two new Truncatellina species from Croatia were described (click
here and search for paper 17). In our study we chose T. rothi from
Greece.
In the internet there are some descriptions of current ideas and research interests
in aeolian sand transport (algorithmic
analysis, electrostatic
effects, wind
erosion problems).
This page is managed by Francisco Welter-Schultes.
Last modified 2.10.2002.
|
Francisco Welter-Schultes |
E-mail:
|
.
KIRCHNER, CH., KRÄTZNER, R. & WELTER-SCHULTES, F. W. (1997): Flying snails - how far can Truncatellina (Pulmonata: Vertiginidae) be blown over the sea? -- Journal of Molluscan Studies 63: 479-487. London. (Abstract) |
.
Here is the text of our Flying-snails paper, with tables
but without most of the figures.
.
FLYING SNAILS - HOW FAR CAN TRUNCATELLINA
(PULMONATA: VERTIGINIDAE) BE BLOWN OVER THE SEA ?
.
CH. KIRCHNER, R. KRÄTZNER1 & F. W. WELTER-SCHULTES2
Max-Planck-Institut für Biophysikalische
Chemie, Abteilung Molekulare Entwicklungsbiologie, Am Fassberg, D-37077 Göttingen,
Germany.
(Current address: Charlotte Kirchner, Max-Planck-Institut
für Züchtungsforschung, Abt. Molekulare Pflanzengenetik, Carl-von-Linné-Weg
10, D-50829 Köln, Germany)
1 Institut für Molekulare Genetik der Universität, Grisebachstr.
8, D-37075 Göttingen, Germany.
2 Zoologisches Institut der Universität, Berliner Str. 28, D-37073
Göttingen, Germany
.
ABSTRACT With populations of land snails of very small size like Vertiginidae,
questions have arisen as to whether populations of relatively distant
islands in archipelagos are really isolated from each other. Apart
from other flight agencies, airborne transport of loose specimens
is not improbable in stormy weather conditions. Currently, mechanisms
of wind-borne transport of sand particles over short and long distances
have been intensively studied. The results are available in the
literature on sediments, allowing the calculation of probable flight
distances for particles in suspension. |
|
.
INTRODUCTION
The smallest pulmonate inhabitants of the Greek islands belong to the genus
Truncatellina (Vertiginidae). A widespread species in the South Aegean
is Truncatellina rothi (Reinhardt) (Fig. 1). The systematics in the Southeast
European Truncatellina has not been thoroughly studied yet. Maybe T.
rothi is a species complex.
The area of dispersal of T. rothi is not well known, presumably due to
its small size. It has been found at several sites in Northern Greece and Albania
(Fig. 2) (Frank, 1987; Klemm, 1962; Maassen, 1984; Dhora & Welter-Schultes,
1996). It is reported to live in the southwestern and southern parts of Turkey
(Schütt, 1993), and is also known from Attikí (Reinhardt, 1916)
and several Greek islands: Lefkáda (Klemm, 1962), Thásos (Reischütz,
1983), Límnos (Reischütz, 1986), Chíos (Bar & Butot,
1986), Náxos (Mylonas, 1982), Léros (Reischütz, 1985), Kálimnos
(Reischütz, 1986), Ródos (Maassen, 1981), Crete, Kíthira
and Andikíthira (Vardinoyannis, 1994) and some surrounding islands of
Crete.
During investigations on the small surrounding islands of Crete by F. Welter-Schultes
in 1987-1994, Truncatellina was found for the first time in ground-litter
samples of Koufonísi Island (South of Crete) in 1991. After 1991, Truncatellina
was found in similar habitats on almost every island investigated (Gávdos,
Gavdopoúla, Chrisí, Grándes, different sites in Crete).
T. rothi was found on the island of Gávdos and in Albania, in
the altitude of 200 m and 400 m respectively.
If there was a probability for Truncatellina to be dispersed by wind
for some kilometers flying from one island to another, the probability of genetic
interchange between island populations would increase. Wind-borne transport
is regarded as an important factor for the dispersal of small species of land
snails. Most of the information available about dispersal ability in land snails
are deductions from distribution patterns (Baur & Bengtsson, 1987). The
relatively quick dispersal of small species northwards and to the tops of mountains
in the Late and Post Glacial Period in Europe is considered to have been wind-borne
(Ant, 1963). There is also strong evidence that the land snail fauna of the
Pacific islands originated primarily through aerial dispersal (Valvolgyi, 1975),
at least for small species.
Mechanisms of airborne dispersal of snails have never been studied under experimental
conditions, as has been the wind-borne transport of sand in desert sand storms
(Anderson, Sørensen & Willets, 1990). The wind-borne translocation
behavior of particles like sand or snow has also been studied under field conditions
(Jensen, Rasmussen, Sørensen & Willetts, 1984; Takeuchi, 1980).
Wind blowing over a surface will, under certain conditions, impart momentum
to any available loose small particles, causing them to skip along the surface.
As each particle impacts the surface, yet more particles are ejected into the
wind, and eventually a distinct layer forms consisting of particles in flight
across the surface. This phenomenon is known as saltation. Very light particles,
for which the force of gravity is small compared to that of aerodynamic origin,
travel downwind at the mercy of turbulent fluctuations without undergoing impact
with the surface. There is no particle-bed-interaction. These particles are
in suspension. We suppose that saltation plays an important role in aerial snail
dispersal on land, in the same way as it does in the case of wind-borne continental
dispersal of sand (Bagnold, 1941; Barndorff-Nielsen, Blæsild, Jensen &
Sørensen, 1983; Sørensen, 1988). Saltation on the sea surface
is impossible for the snails. They have to travel in suspension.
The values of sand, silt and clay when travelling long distances in suspension
are known. A grain of sand (diameter 0.1 mm, fall velocitiy 0.824 m s-1) may
be dispersed by wind (15 m s-1) for 0.3 to 3 s reaching a distance of 46-460
m. For silt grains (diameter 0.01 mm, fall velocity 0.00824 m s-1), a maximum
flight distance in suspension (wind 15 m s-1) of 400-4000 km is calculated (Pettijohn,
Potter & Siever, 1987). For these calculations the sand particle density
is generally assumed to be 2.65 g cm-3 (Iversen & White, 1982).
For calculations of possible flight distances, one of the most important factors
is the fall velocity (Bagnold, 1941). The request of the present study is to
find out the fall velocity of Truncatellina rothi.
MATERIALS AND METHODS
Two random samples of 50 empty shells of Truncatellina rothi from
two different ground litter samples which have been collected on Gávdos
Island (UTM KU3559 and KU3460, for the 1x1 km UTM map of Gávdos see Welter-Schultes,
1995) have been measured (shell height and shell diameter) under microscope.
The shells of the sample KU3460 have been filled with paraffin jelly (Vaseline)
to simulate approximately the live weight of Truncatellina. The weight
of the specimens was determined using an analytical balance of 0.00001 g accuracy
(calculated error ± 20 mg).
Each individual of the two samples was dropped from 5.1 m and from 10.9 m altitude
above base level. The time between dropping, from release to landing was measuered
with a stop watch (accuracy 0.01 s, calculated error of this method ±
0.15 s). The terminal fall velocity has been obtained in evaluating the results
of the experiments.
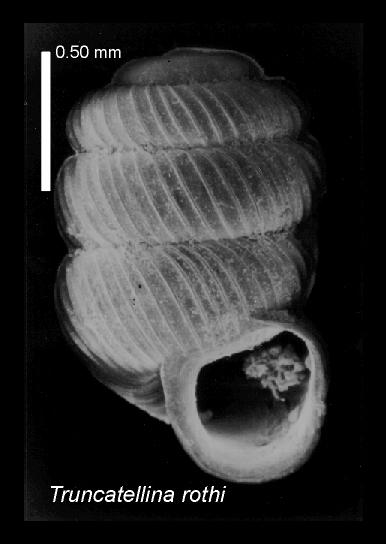
The photo was taken using an SEM Zeiss Novoscan 30 microscope.
RESULTS
Measurements and weight of the specimens
The diameter of the shells (D) is between 0.70 and 0.95 mm (ø = 0.83 mm, sigma x = 0.04 mm, n = 100), the height of the shells (H) varies between 1.1 and 1.8 mm (ø = 1.47 mm, sigma x = 0.12 mm, n = 100) for the specimens of Gávdos (Fig. 3). There is obviously no correlation between diameter and height of the shell of Truncatellina rothi.
Living snails of Truncatellina rothi have not been found on Gávdos.
In 1995, some living individuals of Truncatellina cylindrica (Férussac),
a species which is very similar to T. rothi in size, have been collected in
San Marino. The weight of these snails had an average value of 350-400 micg.
The weight of empty adult shells of Truncatellina rothi was between 100
and 300 micg (ø = 151 micg, sigma x = 34 micg, n = 50). After being filled
with paraffin jelly, the weight of the specimens of sample KU3460 reached values
between 250 and 550 micg (ø = 375 micg, sigma x = 69 micg, n = 50). The
weight differences were principally due to the different size of the shells
and to dust particles in the interior space of the empty shells and on the shell
surface (Fig. 1).
The density of shells filled with Vaseline varied between 0.6 and 0.9 g cm-3
(ø = 0.72
g cm-3, sigma x = 0.14 g cm-3, n = 50, fitting well with
the few values we had for living snails), the density of empty shells 0.2-0.4
g cm-3 (ø = 0.306 g cm-3, sigma x = 0.054 g cm-3,
n = 50). These values have been obtained in dividing the weight of the specimens
by their volume. The approximate volume of Truncatellina can be obtained
by the equation
V = 4/3 × pi × H/2 × (D/2)2 = 0.5236 × H × D × D
V = volume (mm3); H = height of the shell (mm); D = diameter of the shell (mm).
which is the volume of an ellipsoid. For Truncatellina an average
volume of 0.525 mm3 was calculated. The density of living Truncatellina
is lower than that of water because the body of the living snail does not occupy
the entire space inside the shell.
The height of the ribs of Truncatellina rothi is approximate 15 micm,
the mean rib distance varies between 50 and 80 micm.
Fall velocity
The results of the experiments are shown in Fig. 4. The fall velocity values of a grain of sand of the same dimensions as Truncatellina and the values of the empty shells are included in the diagram in order to be compared with the fall velocity values of the "living" snails. The figures show the degree to which living Truncatellina and shells may be expected to vary as regards to their wind resistance. The fall velocity depends on the g cm-2 values of the specimens. For living Truncatellina a mean value of 0.28-0.32 g cm-2 is calculated. The more important factor influencing this value is the weight since differences in shell size are small and can be ignored. Table 1 shows the average terminal fall velocity values for snails of approximate life weight, which have been obtained in the experiments. For living Truncatellina of mean weight, regularly grown and free of any large adherent objects on their shell surface, an average fall velocity of 2.6-2.7 m s-1 was determined.
Theoretical flight distances
The simple addition of the two vectors of the terminal forward velocity, which is assumed to be close to the wind velocity, and the terminal downward velocity, which is assumed to be close to the fall velocity of the snails, results in a theoretical flight distance at laminar wind conditions (Fig. 5B). We base our calculations of flight distances on the assumption that the snail will start from an island from a certain altitude above sea level (100 m). Due to the slower particle response as a result of drag conditions (Anderson, 1987), particles of the size of Truncatellina will not immediately follow the trajectories of the wind turbulences on the lee side of an island. The terminal forward vector as shown in Fig. 5B principally does not describe an unreal situation.
Turbulent wind conditions
Particles in suspension follow two parameters. Suspension is the balance
between downward advective flux as a result of the settling of grains (Table
2), and their upward diffusive flux as a result of turbulence (Anderson &
Hallet, 1986).
As shown in Anderson (1987), it is possible to incorporate turbulent wind conditions
in statistical approaches on trajectories of particles in suspension, allowing
calculations of probable maximum flight distances under natural conditions.
Grains of sand (diameter 0.1 mm, fall velocity 0.824 m s-1) are able
to reach a maximum flight distance of 46-460 m in 15 m s-1 wind.
The maximum height reached by the particles is 0.61-6.1 m respectively (Pettijohn
et al., 1987). The theoretical flight distance (as applied in Table 2, taken
into account the different fall velocity and wind velocity of the given example),
neglecting the upward movement of the grains and setting them to start at an
altitude of 0.61-6.1 m, would be of 11.10-111.0 m.
In our calculations, the upward diffusive flux as a result of turbulent wind
conditions can be included as a turbulence factor, comparing the theoretical
flight distances of sand grains used in the example with their actual maximum
flight distances as given in Pettijohn et al. (1987). This factor 460/111 =
4.14 could be applied to approximate probable maximum flight distances under
natural conditions.
The point at which a particle in saltation is at the top of its trajectory can
be calculated (Sørensen, 1990). Suspension is an extreme kind of modified
saltation, and eolian suspension can be modelled in the same way as modified
saltation (Anderson et al., 1990). The relation between the mean rising periods
and the mean falling periods of the trajectories is 24:76 % (Anderson, 1987).
This relation gives good fits for the suspension profiles and can be applied
for the example in Pettijohn et al. (1987). When only the mean falling periods
of the trajectories are considered the turbulence factor is reduced from 4.14
to 3.15.
The maximum flight distance for one specimen of Truncatellina rothi in
a storm of a wind velocity of 100 km h-1, when starting at an altitude
of 100 m above sea level, is calculated to be approximately 3300 m. If wind
velocity is reduced to 50 km h-1, the distance is halved. Setting
the snails to start at an altitude of 200 m enables them to reach a distance
of 6600 m in 100 km h-1 wind. The probable maximum flight distance
of snails living in 500 m altitude is 16.5 km.
DISCUSSION AND CONCLUSIONS
Outline of the study
The present study has been carried out in order to ascertain whether it is theoretically possible for Truncatellina to overcome distances of several kilometers continuously over sea. The study has not been carried out in order to simulate field conditions, or to calculate exactly possible flight distances of Truncatellina rothi between two Greek islands under natural circumstances. Passive dispersal by wind in general and suspension trajectories in particular are stochastic rather than deterministic. We have also avoided the question of how the snails may be dislodged and lifted into the air. Our studies are thought to give an answer to what is possible once they are airborne.
Neglected influences
Considering the mere fall velocity in our experiments and calculations, we
neglect possible influences originated not only in variation of aerodynamic
forces, but also in the existence of inter-particle forces due to moisture,
electrostatic effects, and other forces of cohesion. These forces are known
to be greater for small particles and relatively independent of particle density
(Iversen, Pollack, Greenley & White, 1976). In comparison to the sand grains
usually dealt with in the sedimentological studies, Truncatellina does
not belong to the small particle fractions.
We also neglect probable changes in wind velocity, and assume that the forward
velocity of the snails equals the velocity of the wind. The wind is faster when
striking over the top of an island (Fig. 5A), but at the same moment the snails
will not yet have reached their terminal forward velocity, due to drag conditions.
Biogeographical implications
Direct passive dispersal by wind is not the only method of airborne translocation
for Truncatellina. There are many other means for living land snails
to be tranported over the sea. Dispersal of land snails by birds and insects
is considered as fact (Rees, 1965; Valvolgyi, 1975). Furthermore, minute snails
are able to stick on leaves, single bird feathers or other inter-island flight
agencies, which can be transported by wind much more easily. In our research,
direct dispersal has been studied because of the presumably increased probability
for single snails to be dislodged by wind.
Initiation of Truncatellina populations does not necessarily require
more than one individual landing on the next island, as Vertiginidae are self-fertile
in many instances (Falkner, 1990). Our results are important concerning probabilities
of genetic interchange, and hence questions of systematics and taxonomy. Relatively
frequent genetic interchange between island populations of minute snail species
are not provided by changing sea levels combined with tectonic movements, though
for explaining the distribution of the Aegean land snails, these events may
be of importance (Heller, 1976).
Flying to islands around Crete
100 km h-1 is probably about the maximum wind velocity in the
Aegean. At this velocity a flight from Andikíthira to Crete or back is
not probable, and Crete could not be reached by Kárpathos snails. So
Crete is isolated from the rest of Greece. The island of Gávdos could
be reached by snails starting their trajectory in 1500 m altitude in Western
Crete. So genetic interchange is possible, but only one-way. Gávdos does
not exceed 400 m altitude, so they are not able to fly back to Crete. Chrisí
and Koufonísi could be reached from various sites from Eastern Crete.
In Eastern Crete the snails would also be able to be transported to Grándes,
Elása and the Dionisádes. The island of Día could probably
be reached by snails starting from the top of Mount Gioúchtas (700-800
m), and from the tops of the mountains of Ródia west of Día.
We base our calculations on the assumption that Truncatellina lives on
the highest points of the islands at the start of the trajectory. Of course,
the greatest distances would not be travelled frequently, but they appear to
be theoretically possible.
ACKNOWLEDGEMENTS
We wish to express our gratitude to W. Zarnack (Göttingen) for helpful comments and kindly placing at our disposal the wind channels of the I. Zoological Institute of Göttingen University, for further experiments which helped to ascertain the results of the presented study. L. Bull (Freetown) and P. Mordan (London) are acknowledged for the linguistic revision and correction of the english manuscript.
REFERENCES
ANDERSON, R. S. 1987. Eolian sediment transport as a stochastic
process: the effects of a fluctuating wind on particle trajectories. Journal
of Geology, 95: 497-512.
ANDERSON, R. S. & HALLET, B. 1986. Sediment transport by
wind: toward a general model. Geological Society of America Bulletin, 97: 523-535.
ANDERSON, R. S., SØRENSEN, M. & WILLETTS, B. B. 1990.
A review of recent progress in our understanding of aeolian sediment transport.
Research Reports, Department of Theoretical Statistics, Institute of Mathematics,
University of Aarhus, 213: 1-48.
ANT, H. 1963. Faunistische, ökologische und tiergeographische
Untersuchungen zur Verbreitung der Landschnecken in Nordwestdeutschland. Abhandlungen
aus dem Landesmuseum für Naturkunde zu Münster in Westfalen, 25: 1-125.
BAGNOLD, R. A. 1941. The physics of blown sand and desert dunes.
Methuen & Co., London.
BAR, Z. & BUTOT, L. J. M. 1986. The land snails of Chios.
De Kreukel, 22: 65-93.
BARNDORFF-NIELSEN, O. E., BLÆSILD, P., JENSEN, J. L. &
SØRENSEN, M. 1983. The fascination of sand. Research Reports, Department
of Theoretical Statistics, Institute of Mathematics, University of Aarhus, 93.
BAUR, B. & BENGTSSON, J. 1987. Colonizing ability in land
snails on Baltic uplift archipelagos. Journal of Biogeography, 14: 329-341.
DHORA, DH. & WELTER-SCHULTES, F. W. 1996. List of species
and atlas of the non-marine molluscs of Albania. Schriften zur Malakozoologie,
9: 90-197.
FALKNER, G. 1990. Binnenmollusken. In: Fechter, R. & Falkner,
G. Weichtiere. Europäische Meeres- und Binnenmollusken. Steinbachs Naturführer,
10: 112-280.
FRANK, CH. 1987. Beitrag zur Kenntnis der Molluskenfauna der
östlichen Mittelmeerländer. Teil III (1): Zusammenfassung der Sammelergebnisse
der Jahre 1982-1985 vom kontinentalen Griechenland, dem Peloponnes, den Nördlichen
Sporaden sowie einigen Inseln des Ionischen und des Ägäischen Meeres.
Malakologische Abhandlungen, 12: 101-124.
HELLER, J. 1976. The biogeography of enid land snails on the
Aegean Islands. Journal of Biogeography, 3: 281-292.
IVERSEN, J. D., POLLACK, J. B., GREENLEY, R. & WHITE, B.
R. 1976. Saltation threshold on Mars: the effect of interparticle force, surface
roughness, and low atmospheric density. Icarus, 29: 381-393.
IVERSEN, J. D. & WHITE, B. R. 1982. Saltation threshold on
Earth, Mars and Venus. Sedimentology, 29: 111-119.
JENSEN, J. L., RASMUSSEN, K. R., SØRENSEN, M. & WILLETTS,
B. B. 1984. The Hanstholm experiment 1982: sand grain saltation on a beach.
Research Reports, Department of Theoretical Statistics, Institute of Mathematics,
University of Aarhus, 125.
KLEMM, W. 1962. X. Teil. Die Gehäuseschnecken. In: Beier,
M. Zoologische Studien in West-Griechenland. Sitzungsberichte, Österreichische
Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse, Abteilung
I, 171: 203-258, Tafeln 1-4.
MAASSEN, W. J. M. 1981. De Molluskenfauna van het griekse eiland
Rhodos. De Kreukel, 17: 21-32, pl. 8-15.
MAASSEN, W. J. M. 1984. Enkele vindplaatsen van mollusken in
Noord Griekenland. De Kreukel, 20: 23-34, pl. 1-10.
MYLONAS, M. A. 1982. Meléti páno sti zoogeografía
ke ikología ton cherséon malakíon ton Kikládon.
[The zoogeography and ecology of the terrestrial molluscs of Cyclades]. Unpublished
Ph. D. Thesis, University of Athens.
PETTIJOHN, F. J., POTTER, P. E. & SIEVER, R. 1987. Sand and
sandstone. Second edition. Springer Verlag, New York.
REES, W. J. 1965. The aerial dispersal of Mollusca. Proceedings
of the Malacological Society of London, 36: 269-282.
REINHARDT, O. 1916. Einige Bemerkungen über Pupa minutissima und Verwandte. Nachrichtsblatt
der Deutschen Malakozoologischen Gesellschaft, 48: 158-167.
REISCHÜTZ, P. L. 1983. Ein Beitrag zur Molluskenfauna der
Insel Thasos (Griechenland). Annalen des Naturhistorischen Museums in Wien,
85B: 133-146, Tafel 1.
REISCHÜTZ, P. L. 1985. Ein Beitrag zur Molluskenfauna von
Léros (Dodekanes, Griechenland). Malakologische Abhandlungen, 11: 17-24.
REISCHÜTZ, P. L. 1986. Beiträge zur Molluskenfauna
der Ägäischen Inseln. Malakologische Abhandlungen, 11: 93-103.
SCHÜTT, H. 1993. Türkische Landschnecken. Christa Hemmen
Verlag, Wiesbaden.
SØRENSEN, M. 1988. Radioactive tracer studies of grain
progress in aeolian sand transport. Research Reports, Department of Theoretical
Statistics, Institute of Mathematics, University of Aarhus, 141.
SØRENSEN, M. 1990. An analytic model of wind-blown sand
transport. Research Reports, Department of Theoretical Statistics, Institute
of Mathematics, University of Aarhus, 214: 1-35.
TAKEUCHI, M. 1980. Vertical profile and horizontal increase of
drift-snow transport. Journal of Glaciology, 26 (94): 481-492.
VAGVOLGYI, J. 1975. Body size, aerial dispersal, and origin of
the Pacific land snail fauna. Systematic Zoology, 24: 465-488.
VARDINOYANNIS, K. 1994. Viogeografía ton cherséon
malakíon sto nótio nisiotikó egeakó tóxo.
[Biogeography of land snails in the South Aegean Island Arc]. Unpublished Ph.
D. Thesis, University of Athens.
WELTER-SCHULTES, F. W. 1995. La végétation de l'île
de Gavdos, la plus méridionale de l'Europe. Influence de facteurs historiques
et humains. Biologia Gallo-hellenica, 21: 189-201.
FIGURE CAPTIONS
Figure 1. Truncatellina rothi, with dust particles in the mouth of the empty shell. Scale bar = 0.20 mm.
Figure 2. Truncatellina rothi has been found at sites in Northern Greece and Albania (dots) and on several Greek islands.
Figure 3. Size and dimensions of the specimens of Truncatellina rothi used in the experiments.
Figure 4. Terminal fall velocity of Truncatellina rothi. A. Relation between fall velocity and the g cm-2 values of the specimens. B. Relation between fall velocity and weight of the specimens. For living Truncatellina of an average weight of 350-400 micg, fall velocity values of 2.4-2.9 m s-1 have been determined.
Figure 5. A. Probable natural wind conditions. The velocity of wind is approximately
reflected in the length of the arrows. B. Addition of the two vectors of wind
velocity and fall velocity. The dashed arrow describes the theoretic trajectory
of the snail neglecting turbulent wind conditions.
Table 1. Mean terminal fall velocity for Truncatellina of approximate life weight.
weight g cm-2
values fall velocity
(± 0.01 g cm-2)
(± 0.3 m s-1)
300 micg 0.27 g cm-2
2.5 m s-1
350 micg 0.29 g cm-2
2.6 m s-1
400 micg 0.31 g cm-2
2.7 m s-1
450 micg 0.33 g cm-2
2.8 m s-1
Table 2. Theoretical flight distances for living Truncatellina rothi, as result of the addition of the two velocity vectors shown in Fig. 5B. Start of the snails 100 m above sea level, laminar horizontal wind 27.8 m s-1.
weight
fall velocity theoretical
flight distance
(± 0.3 m s-1)
(± 93 m)
300 micg 2.5 m s-1
1111 m
350 micg 2.6 m s-1
1068 m
400 micg 2.7 m s-1
1029 m
450 micg 2.8 m s-1
992 m
Table 3. Maximum flight distances for living Truncatellina rothi under turbulent wind conditions. Two different calculations are suggested, the most probable values are assumed to be close to the factor 3.15 values. Start of the snails 100 m above sea level, turbulent wind 27.8 m s-1.
weight
flight distance flight distance
applying
applying
factor 4.14
factor 3.15
(± 385 m)
(± 293 m)
300 micg 4600 m
3500 m
350 micg 4422 m
3364 m
375 micg 4342 m
3304 m
400 micg 4260 m
3241 m
450 micg 4107 m
3125 m
muell@schrottmails.de
hier klicken